Để có thể nói một cách dễ hiểu về hemoglobin (Hb), trước tiên, rất hữu ích khi đối phó với myoglobin (Mb) rất giống với hemoglobin nhưng đơn giản hơn nhiều. Giữa huyết sắc tố và myoglobin có mối quan hệ họ hàng nghiêm ngặt: cả hai đều là protein liên hợp và nhóm giả của chúng (phần không phải protein) là nhóm heme .
Myoglobin là một protein hình cầu bao gồm một chuỗi gồm khoảng một trăm năm mươi axit amin (phụ thuộc vào sinh vật) và trọng lượng phân tử của nó là khoảng 18 Kd.
Như đã nói, nó được ban cho một nhóm heme được chèn vào phần kỵ nước (hoặc lipophilic) của protein, bao gồm các nếp gấp liên quan đến cấu trúc α-helix của protein sợi.
Myoglobin bao gồm chủ yếu là các phân đoạn xoắn ốc, có số lượng tám và hầu như chỉ bao gồm các dư lượng không phân cực (leucine, valine, methionine và phenylalanine) trong khi dư lượng cực thực sự không có (axit aspartic, axit glutamic, lysine và arginine); dư lượng cực duy nhất là hai histidine, đóng vai trò cơ bản trong sự tấn công của oxy đến nhóm heme.
Nhóm heme là nhóm chromophore (hấp thụ trong vùng nhìn thấy) và là nhóm chức năng của myoglobin.
Xem thêm: glycated hemoglobin - hemoglobin trong nước tiểu
Một chút hóa học
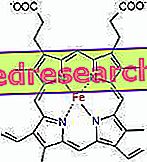
Liên kết giữa protoporphyrin và sắt là một liên kết điển hình của cái gọi là các hợp chất phối hợp là các hợp chất hóa học trong đó một nguyên tử trung tâm (hoặc ion) hình thành liên kết với các loài hóa học khác với số lượng lớn hơn số oxy hóa của nó (điện tích). Trong trường hợp của heme, các trái phiếu này có thể đảo ngược và yếu.
Số phối trí (số liên kết phối trí) của sắt là sáu: có thể có sáu phân tử xung quanh sắt có chung các electron liên kết.
Để tạo thành hợp chất phối trí, cần có hai quỹ đạo với định hướng chính xác: một người có thể "mua" các electron và người còn lại có thể tặng chúng
Trong heme, sắt tạo thành bốn liên kết phẳng với bốn nguyên tử nitơ nằm ở trung tâm của vòng proto-porphyrin và liên kết thứ năm với nitơ của histidine gần; sắt có liên kết thứ sáu của sự phối hợp tự do và có thể liên kết với oxy.
Khi sắt ở dạng ion tự do, các quỹ đạo loại d của nó đều có cùng năng lượng; trong myoglobin, ion sắt liên kết với protoporphyrin và histidine: những loài này phá vỡ từ tính của quỹ đạo d của sắt; mức độ nhiễu loạn sẽ khác nhau đối với các quỹ đạo d khác nhau tùy thuộc vào hướng không gian của chúng và của các loài gây nhiễu. Do tổng năng lượng của các quỹ đạo phải không đổi, nhiễu loạn gây ra sự phân tách năng lượng giữa các quỹ đạo khác nhau: năng lượng thu được từ một số quỹ đạo, tương đương với năng lượng bị mất bởi các quỹ đạo khác.
Nếu sự phân tách xảy ra giữa các quỹ đạo không lớn lắm, thì nên bố trí spin cao điện tử: các electron liên kết cố gắng sắp xếp spin song song trong một số cấp độ phụ có thể (bội số tối đa); mặt khác, nếu nhiễu loạn rất mạnh và có sự phân tách lớn giữa các quỹ đạo, có thể thuận tiện hơn để ghép các electron liên kết trong quỹ đạo năng lượng thấp hơn (độ xoáy thấp).
Khi sắt liên kết với oxy, phân tử sẽ có sự sắp xếp độ xoáy thấp, trong khi đó, khi sắt có liên kết thứ sáu tự do, phân tử sẽ xử lý một spin cao.
Nhờ sự khác biệt về độ xoáy này, thông qua phân tích quang phổ của myoglobin, chúng ta có thể hiểu được liệu oxy có bị ràng buộc (MbO2) hay không (Mb).
Myoglobin là một protein điển hình của cơ bắp (nhưng không chỉ tìm thấy trong cơ bắp).
Myoglobin được chiết xuất từ cá nhà táng, trong đó nó có mặt với số lượng lớn và sau đó được tinh chế.
Cetaceans có hơi thở giống như con người: có phổi phải hấp thụ không khí qua quá trình hô hấp; cá nhà táng phải mang càng nhiều oxy càng tốt vào các cơ có khả năng tích lũy oxy bằng cách liên kết nó với myoglobin có trong chúng; Oxy sau đó được giải phóng chậm khi cetacean được ngâm vì quá trình trao đổi chất cần oxy: lượng oxy mà cá nhà táng có thể hấp thụ càng nhiều, oxy càng có nhiều trong quá trình lặn.

Để làm cho thịt đỏ nhiều hay ít là nội dung của hemoprotein (nó là heme làm cho thịt đỏ).
Hemoglobin có nhiều chất tương tự cấu trúc với myoglobin và có khả năng liên kết oxy phân tử theo cách thuận nghịch; nhưng, trong khi myoglobin bị giới hạn ở các cơ và các mô ngoại biên nói chung, hemoglobin được tìm thấy trong hồng cầu hoặc hồng cầu (chúng là tế bào giả, tức là chúng không phải là tế bào thật) chiếm tới 40% máu.
Hemoglobin là một tetrameter, nghĩa là, nó được tạo thành từ bốn chuỗi polypeptide, mỗi chuỗi có một nhóm heme và giống hệt nhau hai (trong một con người có hai chuỗi alpha và hai chuỗi beta).
Chức năng chính của hemoglobin là vận chuyển oxy ; một chức năng khác của máu trong đó hemoglobin có liên quan là vận chuyển các chất đến các mô.
Trong đường dẫn từ phổi (giàu oxy) đến các mô, hemoglobin mang oxy (đồng thời các chất khác đến mô) trong khi ở đường ngược lại, mang theo chất thải thu được từ các mô, đặc biệt là carbon dioxide được tạo ra trong quá trình trao đổi chất.
Trong sự phát triển của một con người, có những gen chỉ được biểu hiện trong một khoảng thời gian nhất định; Vì lý do này, chúng tôi có các loại huyết sắc tố khác nhau: thai nhi, phôi thai, của người đàn ông trưởng thành.
Các chuỗi tạo nên các loại huyết sắc tố khác nhau này có cấu trúc khác nhau nhưng trên thực tế, chức năng mà chúng thực hiện ít nhiều giống nhau.
Một lời giải thích về sự hiện diện của một số chuỗi khác nhau như sau: trong quá trình tiến hóa của sinh vật, hemoglobin cũng đã phát triển, chuyên vận chuyển oxy từ các khu vực giàu có ở những khu vực thiếu hụt. Khi bắt đầu chuỗi tiến hóa, huyết sắc tố vận chuyển oxy vào các sinh vật nhỏ; trong quá trình tiến hóa, các sinh vật đã đạt đến kích thước lớn hơn, do đó, huyết sắc tố đã thay đổi để có thể vận chuyển oxy ở những khu vực xa hơn so với điểm mà nó giàu có; Để làm điều này, các cấu trúc mới của chuỗi tạo nên huyết sắc tố đã được mã hóa trong quá trình tiến hóa.
Myoglobin liên kết oxy ngay cả ở áp suất khiêm tốn; trong các mô ngoại biên có áp suất (PO2) khoảng 30 mmHg: myoglobin ở áp suất này không giải phóng oxy, do đó sẽ không hiệu quả khi vận chuyển oxy. Mặt khác, Hemoglobin có hành vi đàn hồi hơn: nó liên kết oxy ở áp suất cao và giải phóng nó khi áp suất giảm.
Khi một protein hoạt động chức năng, nó có thể thay đổi hình dạng của nó một chút; ví dụ, myoglobin được oxy hóa có một hình thức khác với myoglobin không oxy và đột biến này không ảnh hưởng đến các loại lân cận.
Diễn ngôn khác nhau trong trường hợp các protein liên quan như hemoglobin: khi một chuỗi bị oxy hóa, nó được tạo ra để thay đổi hình dạng của nó nhưng sự điều chỉnh này là ba chiều và do đó các chuỗi khác của tetrameter cũng bị ảnh hưởng. Thực tế là các chuỗi liên kết lẫn nhau, khiến người ta nghĩ rằng việc sửa đổi một chuỗi ảnh hưởng đến các nước láng giềng khác ngay cả khi ở một mức độ khác; khi một chuỗi được oxy hóa, các chuỗi khác của tetrameter sẽ có "thái độ ít thù địch" hơn với oxy: khó khăn trong việc oxy hóa chuỗi giảm khi các chuỗi gần với nó lần lượt oxy hóa. Bài phát biểu tương tự có giá trị cho khử oxy.
Cấu trúc bậc bốn của deoxyhemoglobin có tên là dạng T (tesa) trong khi cấu trúc của oxyhemoglobin được gọi là dạng R (được giải phóng); ở trạng thái căng thẳng, có một loạt các tương tác tĩnh điện khá mạnh giữa các axit amin và axit amin cơ bản dẫn đến cấu trúc cứng của deoxyhemoglobin (đây là lý do tại sao "dạng căng thẳng"), trong khi khi oxy bị ràng buộc, cường độ của chúng tương tác giảm (đây là lý do tại sao "hình thức phát hành"). Hơn nữa, trong trường hợp không có oxy, điện tích histidine (xem cấu trúc) được ổn định nhờ điện tích trái ngược của axit aspartic trong khi, với sự có mặt của oxy, có một phần protein bị mất một proton; tất cả điều này có nghĩa là hemoglobin oxy hóa là một axit mạnh hơn hemoxybin khử oxy: hiệu ứng bohr .
Tùy thuộc vào độ pH, nhóm heme liên kết dễ dàng hơn với oxy: trong môi trường axit, hemoglobin giải phóng oxy dễ dàng hơn (dạng căng thẳng ổn định) trong khi đó, trong môi trường cơ bản, liên kết với oxy là mạnh hơn

Mỗi huyết sắc tố giải phóng 0, 7 proton trên mỗi mol oxy đến (O2).
Hiệu ứng Bohr cho phép hemoglobin cải thiện khả năng vận chuyển oxy.
Các huyết sắc tố làm cho hành trình từ phổi đến các mô phải tự cân bằng theo áp suất, độ pH và nhiệt độ.
Chúng ta hãy xem ảnh hưởng của nhiệt độ .
Nhiệt độ trong phế nang phổi thấp hơn khoảng 1-1, 5 ° C so với nhiệt độ bên ngoài trong khi ở cơ bắp, nhiệt độ khoảng 36, 5-37 ° C; khi nhiệt độ tăng, hệ số bão hòa giảm (ở cùng áp suất): điều này xảy ra vì nó làm tăng động năng và sự phân ly được ưa chuộng.

Có những yếu tố khác có thể ảnh hưởng đến khả năng liên kết với oxy của hemoglobin, một trong số đó là nồng độ 2, 3 bisphosphoglycerate .
2, 3 bisphosphoglycerate là một chất chuyển hóa có trong hồng cầu ở nồng độ 4-5 mM (không có phần nào khác của cơ thể có mặt ở nồng độ cao như vậy).
Ở pH sinh lý, 2, 3 bisphosphoglycerate là deprotonate và có năm điện tích âm trên nó; nó được kết hợp giữa hai chuỗi beta của huyết sắc tố vì các chuỗi này có nồng độ điện tích dương cao. Các tương tác tĩnh điện giữa các chuỗi beta và 2, 3 bisphosphoglycerate tạo ra độ cứng nhất định cho hệ thống: một cấu trúc căng thẳng thu được có ít ái lực với oxy; trong quá trình oxy hóa, sau đó, 2, 3 bisphosphoglycerate bị trục xuất.
Trong hồng cầu có một bộ máy đặc biệt chuyển đổi 1, 3 bisphosphoglycerate (được sản xuất bởi quá trình chuyển hóa) thành 2, 3 bisphosphoglycerate để nó đạt đến nồng độ 4-5 mM và do đó hemoglobin có thể trao đổi oxy trong các mô.
Huyết sắc tố đến mô ở trạng thái giải phóng (liên kết với oxy), nhưng gần với mô, nó bị carboxyl hóa và chuyển sang trạng thái căng thẳng: protein ở trạng thái này, ít có xu hướng liên kết với oxy, so với trạng thái được giải phóng, do đó hemoglobin giải phóng oxy đến mô; hơn nữa, bằng phản ứng giữa nước và carbon dioxide, có sự sản sinh các ion H +, sau đó thêm oxy cho hiệu ứng bohr.
Carbon dioxide khuếch tán trong hồng cầu đi qua màng plasma; Vì hồng cầu chiếm khoảng 40% máu, chúng ta nên hy vọng rằng chỉ có 40% carbon dioxide lan truyền từ các mô đi vào chúng, thực tế 90% carbon dioxide đi vào hồng cầu vì chúng có chứa một loại enzyme chuyển đổi carbon dioxide trong axit carbonic, kết quả là nồng độ carbon dioxide trong hồng cầu thấp và do đó tốc độ xâm nhập cao.

Một hiện tượng khác xảy ra khi hồng cầu đến mô là như sau: theo độ dốc, HCO3- (dẫn xuất của carbon dioxide) đi ra từ hồng cầu và, để cân bằng đầu ra của điện tích âm, chúng ta có đầu vào clorua gây ra sự gia tăng áp suất thẩm thấu: để bù cho sự thay đổi này, nước cũng được đưa vào gây ra sưng hồng cầu (hiệu ứng HAMCHAER). Hiện tượng ngược lại xảy ra khi hồng cầu đến phế nang phổi: có sự giảm phát của hồng cầu (hiệu ứng HALDane). Do đó hồng cầu tĩnh mạch (hướng đến phổi) tròn hơn so với các động mạch.


